
The effect of probiotic complex of spore-forming bacteria Bacillus on the intestinal microbiome of normal and overweight horses
S. Borovkov, O. Kolchyk, A. Paliy*, V. Borovkova, О. Zlenko, O. Pavlichenko
Sergii BOROVKOV, Candidate of Veterinary Sciences, Clinical Biochemistry Laboratory National Scientific Center ‘‘Institute of Experimental and Clinical Veterinary Medicine’’, Kharkiv, Ukraine; Olena KOLCHYK, Candidate of Veterinary Sciences, Pig Disease Research Laboratory National Scientific Center ‘‘Institute of Experimental and Clinical Veterinary Medicine’’, Kharkiv, Ukraine; Anatoliy PALIY*, Doctor of Veterinary Sciences, Professor, Laboratory of Veterinary Sanitation and Parasitology, National Scientific Center ‘‘Institute of Experimental and Clinical Veterinary Medicine’’, Kharkiv, Ukraine (Corresponding author, e-mail: paliy.dok@gmail.com); Viktoriia BOROVKOVA, Candidate of Veterinary Sciences, Department of Biotechnology, Molecular Biology, and Aquatic Bioresources, State Biotechnological University, Kharkiv, Ukraine. Oksana ZLENKO, PhD in biotechnology, Molecular Diagnostics Laboratory National Scientific Center ‘‘Institute of Experimental and Clinical Veterinary Medicine’’, Kharkiv, Ukraine; Olena PAVLICHENKO, Candidate of Veterinary Sciences, Professor, Doctor of Juridical Science, Department of Sanitation, Hygiene and Forensic Veterinary Medicine, State Biotechnological University, Kharkiv, Ukraine
![]() https://doi.org/10.46419/cvj.57.1.8
https://doi.org/10.46419/cvj.57.1.8
Abstract
Dysbiosis in horses leads to changes in the microbiota composition, fermentation patterns and, ultimately, metabolic disorders. Spore-forming probiotic strains are considered a potential method for restoring normal microbiota and preventing diseases caused by an imbalance of gut microbial pathogens. The aim of the study was to evaluate the effect of a probiotic complex of bacteria of the genus Bacillus on the qualitative and quantitative parameters of the intestinal microbiome in dysbiosis in horses using the 16S sequencing method. The study was conducted on 16 horses (eight healthy working horses and eight overweight animals, both stallions and mares) of different breeds aged 3 to 8 years. It was established that the hindgut microbiome of the studied horses was represented by the phyla Bacillota (ranging from 43.9 to 64.2%), Bacteroidota (to 5.6%), and Pseudomonadota (to 4.8%), with the absence of Actinomycetota, Verrucomicrobiota, and Fusobacteriota. Following the administration of a probiotic complex of Bacillus spp., clinically healthy animals exhibited an increase in the levels of Clostridiaceae and Eubacteriaceae to 7.92% and 23.33%, respectively. Within the Bacillota phylum, the families Staphylococcaceae, Streptococcaceae, and Paenibacillaceae increased to normative levels, ranging from 5.39 to 6.57%, while Peptococcaceae, Veillonellaceae, and Enterococcaceae increased to 2.02–4.74% compared to values before treatment. In overweight animals, a significant increase was observed only to the lower threshold of normative values in Eubacteriaceae (3.17%) and Staphylococcaceae (4.98%), whereas Streptococcaceae increased only to minimal levels (2.88%) relative to pre-treatment values. The relative abundance of Ruminococcaceae was 2.3 times higher in clinically healthy horses compared to overweight animals, whereas prior to treatment, these bacteria were completely absent in the intestines of all studied horses and influenced the overall microbiome balance. Colonisation of the gut microbiome also involved five probiotic Bacillus strains, which were detected in minimal quantities in clinically healthy horses (up to 1.07%) and in overweight animals (up to 0.36%).
Key words: horses; dysbiosis; probiotic; microflora; 16S sequencing; treatment
Introduction
The equine gut microbiome is a complex and diverse ecosystem consisting mainly of bacteria coexisting with viruses, archaea and fungi (Rodriguez et al., 2015; Arroyo et al., 2018; Nishida et al., 2018). As in other mammals, the gut microbiome of horses creates a complex and symbiotic environment that is involved in food digestion, and metabolic functions of protection against pathogens and stimulation of the immune system, directly or indirectly affecting various physiological processes of the host (Shirazi-Beechey, 2008; Barko et al., 2018). The faecal microbiota of horses is in a continuous state of adaptation of their ratios due to changes in feeding, supplementary feed and weather conditions (Dougal et al., 2014; Proudman et al., 2015).
An appropriate and balanced diet is essential for optimal nutrient uptake and health of horses, and improper feeding can cause dysbiosis and a decrease in overall resistance (Costa and Weese, 2012; Warzecha et al., 2017; Borovkov et al., 2024). Dysbiosis (Blackmore et al., 2013; Venable et al., 2016; Salem et al., 2018) is associated with feeding a high concentrate diet, poor quality feed, confinement, stress, hunger, age, and medical interventions. In recent years, some authors have linked changes in the gut microbiome of horses to colic, as they have observed changes in the number of certain groups of bacteria that play an important role in maintaining gastrointestinal health, leading to changes in colon pH and production of volatile fatty acids (Costa and Weese, 2018; Blikslager, 2019; Salem et al., 2019).
However, antibiotic therapy often complicates gut dysbiosis and polymicrobial infections (Boucher et al., 2024). In some cases, antibiotics can cause the development of multidrug-resistant mutants (Hadzevych et al., 2022). In particular, sublethal levels of antibiotics improve the production of virulence factors and increase bacterial resistance in epithelial cells (Liu et al., 2020; Ghuneim et al., 2022). The use of spore-forming probiotics of Bacillus bacteria, which have the ability to sporulate abundance sporulate to survive in the intestinal lumen and eliminate invasive pathogens, is promising. The antimicrobial peptides secreted by Bacillus bacteria have a huge advantage over traditional antibiotics, as they are similar to the antimicrobial peptides secreted as part of the animal’s natural immunity (Kolchyk et al., 2022). In addition, Bacillus probiotics produce a number of nutrients, including extracellular polysaccharides, vitamins and exoenzymes, which promote the growth of beneficial microbiota. The extracellular polysaccharides produced by Bacillus probiotics can serve as a carbon source for lactobacilli and enhance their ability to adhere and produce acetate (Cai et al., 2020). The use of spore-forming probiotics can reduce the spread of pathogens in animals, improve digestion and absorption of nutrients in the intestine, and maintain a healthy microecological balance.
The aim of this study was to evaluate the effect of the probiotic complex of bacteria of the genus Bacillus on the qualitative and quantitative indicators of the horse intestinal microbiome.
Materials and Methods
Clinical examination of animals and sampling was carried out in 2024 at the equestrian sports complex of the State Biotechnological University, Mala Danylivka village, Kharkiv district, Kharkiv region, Ukraine. Faecal samples were collected from 16 horses (8 working healthy horses and 8 overweight animals, both stallions and mares) of predominantly Ukrainian horse breeds aged 3 to 8 years: 9 animals of the Ukrainian Warmblood breed, 3 animals of the Draft Horse breed, and 4 outbred animals. The physical condition of the horses was assessed using the Body Condition Score (BCS) scale, based on a visual inspection and palpation of key body areas: the neck, withers, ribs, loin, tailhead, and shoulders. The evaluation was performed by a veterinary physician, who assigned scores ranging from 1 (severely emaciated) to 9 (obese). Horses were considered overweight at a BCS of 7 or higher. An average score was calculated for each horse, reflecting the level of body fat and overall physical condition. The average body weight of clinically healthy horses was 504±12.4 kg (range 482–511 kg) and that of overweight horses was 523±23.2 kg (range 490–580 kg). Faecal samples (each 50 g) were collected from the rectum of each animal under aseptic conditions by hand using sterile rubber gloves and transferred into special plastic containers, which were immediately frozen in liquid nitrogen for further transport. The diets of the 16 experimental animals prior to addition of the probiotic complex of bacteria of the genus Bacillus in clinically healthy and overweight horses did not differ in composition and included: grass hay 9 kg, straw 3–4 kg, oats 2 kg, barley 300 g, corn 200 g, table salt 29 g. All horses were housed in the same facility, in individual stalls.
The probiotic complex of bacteria of the genus Bacillus was administered to experimental animals orally at a dose of 20 billion CFU/mL, mixed with 400 g feed in the morning for 21 days. Horses from both groups were put out to pasture together. Animals had free access to clean water. Animals were dewormed with Albendazole-L 10% 2 weeks before the experiment and no antimicrobial drugs were used. The clinical condition of the horses was monitored daily with an assessment of their general condition, including faecal consistency and body temperature. Faecal samples for 16S sequencing were collected prior to the start of and after 3 weeks of administration of the probiotic complex.
The probiotic complex of bacteria of the genus Bacillus includes five spore-forming strains: Bacillus subtilis UNCSM – 020, Bacillus licheniformis UNCSM – 033, Bacillus amyloliquefaciens ALB – 65, Bacillus pumilus UNCSM – 026, and Bacillus subtilis variant mesentericus UNCSM – 031.
Laboratory studies were conducted at the National Scientific Centre Institute of Experimental and Clinical Veterinary Medicine and the Institute of Veterinary Medicine of the NAAS.
Probiotic spore-forming strains were cultured on standard nutrient media, i.e., meat-peptone broth (MPB), meat-peptone agar (MPA) with 5 g yeast extract and 0.1% glucose at pH 6.8–7.0. Each of the five probiotic strains was cultivated separately in 3 L flasks without shaking for 5 days at 37.0±0.5°C. The culture liquids obtained in this way were sterilely combined in a 1:1 ratio by volume. Total DNA was extracted from faecal samples using a PureLink Microbiome DNA purification kit (Invitrogen, USA, cat. no. A29789) according to manufacturer instructions.
To prepare the libraries, the 16S barcoding kit 1-24 (Cat. No. SQK16S024, Oxford Nanopore, USA) was used to amplify specific regions of the 16S RNA gene and to barcode the samples. To purify the libraries, NucleoMag NGS Clean-up and Size select magnetic beads (ref 744970.50, lot 2311-002, Macherey-Nagel, Germany) were used according to the recommended protocol Rapid sequencing amplicons – 16S barcoding (SQK- 16S024).
Metagenomic sequencing by NGS Nanopore technology (Oxford Nanopore, USA) was performed using the ONT MinION instrument (Oxford Nanopore, USA), using the Flow Cell Priming Kit (EXP-FLP002, Oxford Nanopore, USA) and the Spot-on Flow Cell R9.4.1 flow cell (FLO-MIN106D, Oxford Nanopore, USA) according to the standard protocol recommended by Oxford Nanopore, USA.
The DNA concentration at all stages of the study was determined on a Qubit v.3 fluorometer (Thermo Fisher Scientific, USA) using a Qubit 1xdsDNA HS assay kit (Invitrogen, USA), and DNA purity was determined on a Nanodrop spectrophotometer (Thermo Fisher Scientific, USA) by the absorbance ratio at wavelengths of 260/280 (at least 1.8) and 260/230 (at least 1.7). The library was diluted at equimolar concentrations and used for sequencing at a concentration of 70 fmol per 10 μL 10 mM Tris HCl pH 8.0 with 50 mM NaCl. For 16S amplicons of ~1500 bp, 50–100 fmol corresponds to ~50–100 ng DNA.
The raw data were baselined using the Guppy 6.1.7 software package with the ultra-high accuracy model (dna_r9.4.1_450bps_sup.cfg) and the filtering of reads with quality below 10. The guppy_barcoder utility of this package demultiplexed the data with the removal of barcodes. The average read quality was 13.5, with a median of 12. The average read length was 1190, with a median of 1431.
The taxonomic classification of the 16s gene was performed in the 16s workflow of EPI2ME Desktop 24.02-01 software. Minimap2 was used as a classifier based on the ncbi_16s_18s database.
Statistical analysis of the data was carried out using Minitab 19, Minitab Inc. in a free trial version. Based on the results of statistical processing, the tables showed non-parametric indicators, such as: Median and Standard Deviation in the format Mean±StDev, and a significant difference between healthy and overweight horses was established based on the Mann Whitney U test (P<0.05).
During the experimental studies presented here, all work with horses involved in the study was performed in compliance with the basic bioethics principles, pursuant to Article 26 of the Law of Ukraine ‘On Protection of Animals from Cruelty’, the European Convention for the Protection of Vertebrates Used for Experimental and Other Scientific Purposes (1986), and the General Ethical Principles for Animal Experiments adopted by the First National Congress on Bioethics (2012).
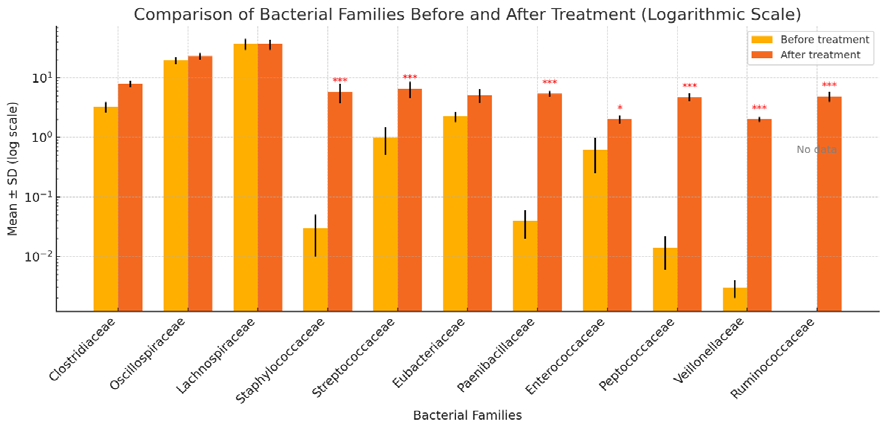
Figure 1. Relative frequency of the main families of the phylum Bacillota in clinically healthy horses prior to and after the use of a probiotic complex of bacteria of the genus Bacillus (*P<0.05; ** P<0.01; *** P<0.001 compared to the levels prior to treatment)
Results and Discussion
In adult horses, the intestinal microbiome can be unstable and is influenced by age, breed, feeding, environmental conditions (access to pasture, diet, season), and the use of probiotics and antimicrobials. All of these factors can lead to changes in the fermentation pattern of the gut microbiome and ultimately to metabolic disorders.
The distribution and relative frequency of bacterial species in the hindgut of 16 experimental horses before the experiment, as confirmed by 16S sequencing, showed that Bacillota was the dominant phylum in the taxonomic structure, with 10 families present in abundance ranging from 43.9 to 64.2%, exceeding the number of Bacteroidota species up to 5.6%, and Pseudomonadota up to 4.8%, with the absence of Actinomycetota, Verrucomicrobiota and Fusobacteriota.
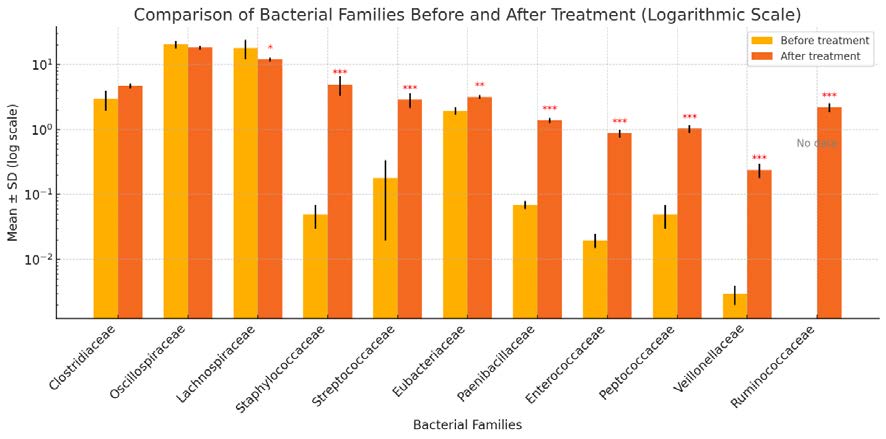
Significant shifts at the level of phylum Bacillota in the intestinal microbiome occurred in all 16 horses. In the eight clinically healthy horses, after oral administration of the probiotic complex of bacteria of the genus Bacillus, representatives of the order Clostridiales and the families Oscillospiraceae and Lachnospiraceae were recorded at abundances of 23.33 and 37.04%, respectively, with no significant changes, while in overweight horses a decrease of 6% in Lachnospiraceae was recorded (Figures 1 and 2).
In clinically healthy animals, the families Clostridiaceae and Eubacteriaceae increased by 2.4 and 2.3 times after administration of the probiotic complex of bacteria of the genus Bacillus, as compared to a 1.6 times increase in overweight horses.
Facultative anaerobic bacteria of the families Staphylococcaceae (5.85%), Streptococcaceae (6.57%), Paenibacillaceae (5.39; P<0.001), Enterococcaceae (2.02%; P<0.05), Peptococcaceae (4.74%), Veillonellaceae (2.02; P<0.001) were found in relatively low numbers in the hindgut after treatment, compared to their levels before treatment (Figure 1).
In overweight animals, a significant increase was recorded only for the families Staphylococcaceae (to 4.98%), Streptococcaceae (2.88%; P<0.001) and Eubacteriaceae (3.17%; P<0.01), respectively, compared to the values prior to treatment. The levels of the families Veillonellaceae (0.24%), Enterococceae (0.88%; P<0.001) did not exceed the lower limit of the minimum values (Figure 2).
Representatives of the family Ruminococcaceae increased to 4.93% in clinically healthy horses and to 2.19% (P<0.001) in overweight animals, while before treatment, these bacteria were not detected in the basic microbiota of any of the 16 horses (Figures 1 and 2).
The probiotic bacteria Bacillus of the family Bacillaceae colonised the gastrointestinal tract and changed the gut microbiome of animals, and the transition between the dormant spore and vegetative cell in Bacillus contributed to greater persistence in the intestinal microbiome. After three weeks of treatment, four strains of Bacillus were detected in minimal amounts in clinically healthy horses from 0.56 to 1.07% and from 0.02 to 0.36% in overweight animals (Table 1). Dipicolinic acid present in the spores of probiotic Bacillus strains inhibits in vitro growth of most lactobacilli and enterobacteria, and activates both digestive and metabolic processes.
Figure 2. Relative frequency of the main families of the phylum Bacillota in overweight horses prior to and after the use of a probiotic complex of bacteria of the genus Bacillus (*P<0.05; ** P<0.01; *** P<0.001 compared to values before treatment)
The phylum Pseudomonadota accounted for 4.7 and 2.0% of the total gut microbiome and was represented by low diversity in both healthy and overweight animals of the Enterobacteriaceae family with two genera (Escherichia 1.9 and 0.06%, and Shigella 2.53 and 1.10%; p<0.05, respectively), and the families Pseudomonadaceae (0.28 and 0.05%; p<0.001), and Neisseriaceae (0.03 and 0.85%; p<0.001), respectively. The addition of a probiotic complex of bacteria of the genus Bacillus to animals contributed to the appearance of the genus Salmonella in 16 experimental horses in the normative range from 1.61 to 1.88% (p<0.001), a significant increase in bacteria of the genus Escherichia by 2.1 times (P<0.05) in healthy horses, and Shigella by 2.3 times (P<0.05) and representatives of the family Pseudomonadaceae to 3.37% (P<0.001) in overweight horses (Table 1).
In both healthy and overweight animals, a significantly low composition of representatives of the phylum Bacteroidota and family Flavobacteriaceae was noted (0.55 and 0.03%), which statistically increased to 2.62% (P<0.01) in healthy animals, but did not reach the lower limit of the norm (3.0%), while in overweight animals it was noted in a significantly low amount of 0.37% after treatment. The family Prevotellaceae was within the normative values (14.2%) after oral administration of a probiotic complex of bacteria of the genus Bacillus in healthy animals, and at the lower limit of the norm (3.25%) in overweight animals.In the present study, the phyla Spirochaetota and Actinomycetota were absent in all horses before treatment, while after the inclusion of the probiotic complex in the diet, a statistically significant increase in the phylum Spirochaetota (family Spirochaetaceae and families Borrelia and Brachyspira was noted in clinically healthy horses to 3.98 and 5.44% (p<0.001), respectively, which was 2.3 and 1.7 (P<0.001) times higher than in overweight horses. The phylum Actinomycetota in overweight horses with representatives of the Lactobacillaceae family was 0.80% (P<0.001) and did not differ statistically from healthy horses, while the number of Bifidobacteriaceae bacteria increased 1.6 times (P<0.001) compared to healthy animals after treatment.
Dysbiosis with manifestations of microbiome imbalance and loss of certain bacterial phyla in healthy and overweight working horses against a background of unbalanced feeding increases overall vulnerability, which can lead to colitis.
After administration to healthy and overweight experimental working horses, the probiotic complex of bacteria of the genus Bacillus affected the overall balance of the microbiome of animals, supported the growth of beneficial microorganisms in the intestine with a limited nutrient content and, through direct interaction with opportunistic bacteria using structurally diverse metabolites, contributed to the stability and homeostasis of the intestinal flora.
Animal feeding is a major factor to consider when assessing the gut microbiota. Abrupt changes in diet can lead to a disruption in the stability of gastrointestinal commensal bacteria (Li et al., 2018). The gastrointestinal microbial communities that colonise the colon play an important role in feed digestion, provide protection against pathogen invasion, maintain intestinal epithelial integrity, and promote immune responses (Schoster et al, 2016; Kauter et al, 2019; Paliy et al., 2020).
Table 1. Diversity of the colonic bacterial phyla in clinically healthy (n=8) and overweight horses (n=8) before and after treatment with probiotic complex of bacteria of the genus Bacillus, expressed as Mean±SD
|
Phylum, family/genus |
Clinically healthy horses |
Overweight horses |
||
|
Before treatment |
After treatment |
Before treatment |
After treatment |
|
| Bacillota, Bacillaceae, Bacillus subtilis |
absent |
1.07±0.11 ••• |
absent |
0.36±0.05***,••• |
| Bacillus licheniformis | absent | 0.56±0.14 ••• | absent | 0.02±0.005***,••• |
| Bacillus pumilus | absent | 0.91±0.10 ••• | absent | 0.05±0.01***,••• |
| Bacillus amyloliquefaciens | absent | 0.80±0.13 ••• | absent | 0.34±0.28 ••• |
| Pseudomonadota, Enterobacteriaceae, Escherichia |
1.91±0.64 |
3.98±0.71 • |
0.06±0.03*** |
0.72±0.09***,••• |
| Salmonella | absent | 1.88±0.61 ••• | absent | 1.61±0.62 ••• |
| Shigella | 2.53±0.30 | 1.83±0.56 | 1.10±0.18** | 2.53±0.59 • |
| Pseudomonadaceae | 0.28±0.15 | 0.71±0.10 | 0.05±0.02*** | 3.37±1.42 *** |
| Neisseriaceae | 0.03±0.01 | 0.57±0.11 ••• | 0.85±0.08*** | 0.59±0.14 |
| Bacteroidota, Prevotellaceae | 5.04±0.29 | 14.2±3.15 • | 2.64±0.36*** | 3.25±0.98*** |
| Flavobacteriaceae | 0.55±0.18 | 2.62±0.52 •• | 0.03±0.01*** | 0.37±0.17***,••• |
| Spirochaetota, Spirochaetaceae, Borrelia |
absent |
3.98±0.55 ••• |
absent |
1.75±0.17**,••• |
| Brachyspira | absent | 5.44±0.60 ••• | absent | 3.14±0.49*,••• |
| Actinomycetota. Bifidobacteriaceae | absent | 1.83±0.22 ••• | absent | 2.99±0.26**,••• |
| Lactobacillaceae | absent | 0.73±0.12 ••• | absent | 0.80±0.19 ••• |
Note: * P<0.05; ** P<0.01; *** P<0.001 compared to same group prior to treatment; • Р<0.05; •• Р<0.01; ••• Р<0.001 compared between groups.
The results show that in 16 experimental horses, both clinically healthy and overweight animals showed a significant shift of the intestinal microbiota ratio (dysbiosis). Dysbiosis was manifested in animals by the absence of Fibrobacterota and members of the Ruminococcaceae family, which, together with bacteria of the Lachnospiraceae family, which were found in sufficient numbers, and are able to use complex carbohydrates, including cellulose, for short-chain fatty acids for the production of intestinal epithelial mucus. Lachnospiraceae are prolific producers of butyrate, and butyrate is known to have a protective function for colonocytes in the intestinal wall (Jalanka-Tuovinen et al., 2011; Faubladier et al., 2013). The phylum Verrucomicrobiota, which maintains the integrity of the mucin layer and reduces intestinal inflammation, was also not detected in the experimental animals (Everard et al., 2013).
The families Clostridiaceae and Eubacteriaceae are involved in the digestion of complex and simple carbohydrates, and were sparse in the experimental horses, averaging only 3.36 and 2.24% in healthy horses, and 2.99 and 1.95% in overweight animals, respectively.
Facultative anaerobic bacteria of the families Staphylococcaceae, Streptococcaceae, Paenibacillaceae, Enterococcaceae, Peptococcaceae, Veillonellaceae of the phylum Bacillota were found in relatively low numbers in the hindgut of both healthy and overweight animals. The presence of these bacteria, which consume oxygen present in the intestinal lumen, creates an anaerobic environment that allows the growth of strict anaerobes such as Bacteroidota and Clostridiaceae, thereby maintaining their optimal ratios (Dougal et al., 2014; Venable et al., 2016). Thus, bacteria of the family Flavobacteriaceae of the phylum Bacteroidota were found in low amounts in the faeces of the large intestine of all experimental animals (0.03 in healthy and 0.55% in overweight animals). Representatives of the phylum Bacteroidota use starch, xylan, and pectin, and are among the main degraders of many complex polysaccharides in plant cell walls (Kibegwa et al., 2023).
Proteolytic bacteria of the phylum Pseudomonadota in the hindgut microbiome were low in healthy horses (4.7%) and overweight animals (2.0%), with the absence of representatives of the phylum Spirochaetota and typical intestinal microorganisms of the families Bifidobacteriaceae and Lactobacillaceae in all 16 experimental horses. After all, these bacteria protect the animal body from the reproduction of pathogens, show immunomodulatory activity, and participate in the synthesis of vitamins and essential amino acids (Ishizaka et al, 2014; Gotić et al., 2017).
Dysbioses of the intestinal microbiota in the tested horses depend on feeding, namely an unbalanced diet that does not take into account age, breed, physiological condition, and workload. Hay should be given at 2.0% of the animal’s body weight, straw up to 10 kg, but all animals received the same amount. For overweight horses, succulent feed should be added to maintain the minimum nutrient intake of feed: fodder beetroot 8 kg, silage up to 15 kg, with the complete exclusion of concentrated feed. On the contrary, workhorses with an average workload should be fed green grass to their heart’s content, concentrates 3 kg, hay 2 kg, salt 40 g. When lacking energy and nutrients in the diets, horses tire more quickly and their performance decreases, which leads to shifts in the microbiome and increases the risk of colic.
It is known that the horse gastrointestinal tract is inextricably linked to the immune system (Jensen et al., 2010). In fact, about 70% of immune cells are located in the mucosal membrane of the gastrointestinal tract and lymphoid tissue associated with the gut (Ruemmele et al., 2009). Oral spore-forming probiotic strains such as B. subtilis, B. pumilus and B. licheniforms affect both the innate and adaptive immune system by interacting with different immune cells located in these tissues, and balancing the microbiota composition in the gut (Bernardeau et al., 2017).
The use of a probiotic complex of bacteria of the genus Bacillus in 16 experimental horses demonstrated a positive effect on the intestinal microbiota. The five probiotic strains contained within the composition synthesised polypeptides that were antagonistic to intestinal microorganisms, thereby significantly increasing the digestibility of animal feed. Spore-forming strains of B. licheniformis, B. amyloliquefaciens, B. pumilus, and B. subtilis variant mesentericus maintained the ecological balance of the intestine by consuming oxygen and contributing to the creation of an anaerobic environment, which in turn promoted the growth of dominant bacteria in the intestine.
In healthy working horses and overweight animals, changes occurred in the ratio of the main types of microbiota. The abundance of the phylum Bacillota increased at the expense of representatives of the families Staphylococcaceae, Streptococcaceae, Paenibacillaceae, Enterococcaceae, Peptococcaceae, Veillonellaceae in healthy animals (2.02–5.85%) and in overweight horses (0.24– 4.88%). The occurrence of Ruminococcaceae bacteria in all horses was noted in the range of 2.19– 4.93%. The phylum Spirochaetota in overweight and healthy animals was absent before treatment with the probiotic complex, when it achieved the level of normative values of 4.89 and 9.42%, respectively, and the phylum Actinomycetota (Bifidobacteriaceae and Lactobacillaceae) achieved reached values of 3.79 and 2.56%, respectively. The extracellular polysaccharides produced by Bacillus can serve as a carbon source for lactobacilli, enhancing their ability to adhere and produce acetate. The organic acids produced by Bacillus acidify the gut environment, thereby promoting the enrichment of beneficial bacteria (Cai et al., 2020). The bacteria of the phylum Pseudomonadota in healthy and overweight animals did not differ (8.82 vs 8.97%), while the concentration of Bacteroidota representatives was 3.62 and 16.8%, respectively.
The representation in the hindgut of horses of bacteria of the genus Bacillus (B. licheniformis, B. amyloliquefaciens, B. pumilus, B. subtilis) microorganisms with broad antagonistic properties was low and did not exceed 1.07 and 0.36% in healthy and overweight horses.
Most studies suggest that probiotic bacteria survive and colonise the gastrointestinal tract of horses even after discontinuation of supplementation (Kauter et al., 2019) and may limit the use of new probiotic strains, thereby having a significant impact on the gut barrier through direct interaction with enterocytes (Azad et al., 2018). Increased intestinal permeability in dysbiosis can lead to disease, as the intestinal barrier is no longer able to protect against microbial toxins and pathogens present in the gastrointestinal lumen.
It is known that the simultaneous use of a combination of B. subtilis and B. licheniformis strains improves nutrient absorption and overall digestion (Wang et al., 2021). Another study using the same combination of probiotics found that they secrete protease, lactase, lipase, and amylase enzymes that also provide digestive benefits (Yang et al., 2021). Moreover, Chen and Yu (2020) found an increase in Lactobacillus and Firmicutes when working individually with microbiome diversity, which aligns with our findings after using a Bacillus probiotic complex containing five probiotic strains.
- subtilis has been shown to promote the colonisation of the gut of piglets with beneficial bacteria, including Lactobacillus, Leucobacter, Bifidobacteria, Megasphaera, Coprococcus and Prevotella (Ding et al., 2021; Wang et al., 2021). The addition of B. subtilis strain in the diet increased the number of Lactobacillus species and decreased the number of Clostridium perfringens and E. coli in the colon of sows (Baker et al., 2013).
Similarly, oral administration of B. subtilis ATCC 6051 strain resulted in an increase in Enterococcus in broilers (Ciurescu et al., 2020), and Lactobacillus spp. and Bifidobacterium spp. bacteria in laying hens (Forte et al., 2016). As a result of sequential selection, the strains L. plantarum 7, L. casei 27, L. plantarum 7-317, B. adolescentis 17 and B. adolescentis 17-316 were selected according to their antagonistic and adhesive properties, which are promising for the development on their basis of the domestic probiotic for animals and poultry (Gujvinska and Paliy, 2018).
Thus, the experimental combination of five strains of Bacillus subtilis UNCSM – 020, Bacillus licheniformis UNCSM – 033, Bacillus amyloliquefaciens ALB – 65, Bacillus pumilus UNCSM – 026, Bacillus subtilis variant mesentericus UNCSM – 031 in the probiotic complex had a synergistic potential to restore the ratio of beneficial microbiome due to the release of antimicrobial substances (bacteriocins) that inhibit the development of pathogenic microflora and creates conditions for filling the vacant niche with normal bacteria in the intestine of both healthy and overweight horses.
Conclusions
Dysbiosis in clinically healthy and overweight horses was manifested by a less diverse population and changes in the relative diversity of several phyla Bacillota, Pseudomonadota, Bacteroidota, the absence of Spirochaetota, Actinomycetota, Fibrobacterota, Verrucomicrobiota and the bacterial families Bacillaceae, Ruminococcaceae.
The probiotic complex of bacteria of the genus Bacillus in horses of varying physiological condition confirmed its effectiveness for dysbiosis by increasing abundance and stabilising the microbial diversity of the phyla Bacillota, Pseudomonadota, Bacteroidota, Spirochaetota, Actinomycetota, and by enhancing the intestinal barrier function, preventing the growth of pathogenic bacteria and the risk of colitis in animals.
The probiotic complex of bacteria of the genus Bacillus is useful in the case of digestive disorders associated with unbalanced feeding, dietary changes, and prevention of gastrointestinal diseases in horses, with a high ratio of effectiveness and safety.
References [… show]
Učinak probiotičkog kompleksa sporogenih bakterija Bacillus na crijevni mikrobiom normalnih i pretilih konja
Sergii BOROVKOV, Candidate of Veterinary Sciences, Clinical Biochemistry Laboratory National Scientific Center ‘‘Institute of Experimental and Clinical Veterinary Medicine’’, Kharkiv, Ukraine; Olena KOLCHYK, Candidate of Veterinary Sciences, Pig Disease Research Laboratory National Scientific Center ‘‘Institute of Experimental and Clinical Veterinary Medicine’’, Kharkiv, Ukraine; Anatoliy PALIY, Doctor of Veterinary Sciences, Professor, Laboratory of Veterinary Sanitation and Parasitology, National Scientific Center ‘‘Institute of Experimental and Clinical Veterinary Medicine’’, Kharkiv, Ukraine; Viktoriia BOROVKOVA, Candidate of Veterinary Sciences, Department of Biotechnology, Molecular Biology, and Aquatic Bioresources, State Biotechnological University, Kharkiv, Ukraine. Oksana ZLENKO, PhD in biotechnology, Molecular Diagnostics Laboratory National Scientific Center ‘‘Institute of Experimental and Clinical Veterinary Medicine’’, Kharkiv, Ukraine; Olena PAVLICHENKO, Candidate of Veterinary Sciences, Professor, Doctor of Juridical Science, Department of Sanitation, Hygiene and Forensic Veterinary Medicine, State Biotechnological University, Kharkiv, Ukraine.
Disbioza u konja dovodi do promjena u sastavu mikrobiote, obrazaca fermentacije i naposljetku do metaboličkih poremećaja. Sporogene varijante probiotike smatraju se potencijalnim metodama za ponovnu uspostavu normalne mikrobiote i sprečavanju bolesti prouzročenih neravnotežom crijevnih mikrobnih patogena. Cilj je istraživanja procjena učinka probiotičkog kompleksa bakterija roda Bacillus na kvalitativne i kvantitativne parametre crijevnog mikrobioma kod disbioze u konja, upotrebom metode sekvenciranja 16S. Istraživanje je provedeno na 16 konja (pastusi i kobile) različitih pasmina starosti od 3 do 8 godina (8 zdravih radnih konja i 8 životinja prekomjerne težine). Ustanovljeno je da je mikrobiom debelog crijeva konja u istraživanju zastupljen koljenima Bacillota (od 43,9 % do 64,2 %), Bacteroidata (do 5,6 %) i Pseudomonadota (do 4,8 %), a bez prisutnosti koljena Actinomycetota, Verrucomicrobiota i Fusobacteriota. Nakon uvođenja probiotičkog kompleksa Bacillus spp., klinički zdrave životinje iskazale su povišenje razine Clostridiaceae i Eubacteraceae do 7,92 %, odnosno 23,33 %. Unutar koljenja Bacillota, porodice Staphylococcaceae, Streptococcaceae i Paenibacillaceae povisile su se na normativne razine, u rasponu od 5,39% do 6,57 %, dok su se Peptococcaceae, Veillonellaceae i Enterococcaceae povisile do 2,02% do 4.74 % u usporedbi s vrijednostima prije liječenja. Kod životinja prekomjerne težine značajan je porast primijećen samo do donjeg praga normativnih vrijednosti kod Eubacteriaceae (3,17 %) i Staphylococcaceae (4,98 %), dok su se Streptococcaceae povisile do minimalnih razina (2,88 %) u usporedbi s vrijednostima prije liječenja. Kod klinički zdravih konja u usporedbi s pretilim životinjama, relativno brojnost Ruminococcaceae je bio 2,3 puta veća, dok su prije liječenja te bakterije bile potpuno odsutne u crijevima svih konja u istraživanju te su utjecale na opću ravnotežu mikrobioma. Kolonizacija crijevnog mikrobioma također je obuhvaćala pet probiotičkih varijanti Bacillusa, koji su otkriveni u minimalnim količinama kod klinički zdravih konja (do 1,07 %) i kod pretilih životinja (do 0,36 %).
Ključne riječi: konji, disbioza, probiotik, mikroflora, sekvenciranje 16S, liječenje